Различия раст животн и бактериальных клеток. Сходство и различия строения клеток растений и животных
Общее в строении растительных и животных клеток: клетка живая, растет, делится. протекает обмен веществ.
И в растительных, и в животных клетках имеется ядро, цитоплазма, эндоплазматическая сеть, митохондрии, рибосомы, аппарат Гольджи.
Различия между растительными и животными клетками возникли из-за разных путей развития, питания, возможности самостоятельного движения у животных и относительной неподвижности растений.
Клеточная стенка у растений есть (из целлюлозы)
у животных - нет. Клеточная стенка придает растениям дополнительную жесткость и защищает от потерь воды.
Вакуоль есть у растений, у животных - нет.
Хлоропласты есть только у растений, в которых образуются органические вещества из неорганических с поглощением энергии. Животные потребляют готовые органические вещества, которые получают с пищей.
Резервный полисахарид: у растений – крахмал, у животных – гликоген.
Вопрос 10 (Как организован наследственный материал у про- и эукариот?):
а) локализация (в прокариотической клетке – в цитоплазме, в эукариотической клетке – ядро и полуавтономные органоиды: митохондрии и пластиды), б) характеристика Геном в прокариотической клетке: 1 кольцевидная хромосома – нуклеоид, состоящая из молекулы ДНК (укладка в виде петель) и негистоновых белков, и фрагменты – плазмиды – внехромосомные генетические элементы. Геном в эукариотической клетке – хромосомы, состоящие из молекулы ДНК и гистоновых белков.
Вопрос 11 (Что такое ген и какова его структура?):
Ген (от греч. génos - род, происхождение), элементарная единица наследственности, представляющая отрезок молекулы дезоксирибонуклеиновой кислоты - ДНК (у некоторых вирусов - рибонуклеиновой кислоты - РНК). Каждый Г. определяет строение одного из белков живой клетки и тем самым участвует в формировании признака или свойства организма.
Вопрос 12 (Что такое генетический код, его свойства?):
Генети́ческий код - свойственный всем живым организмам способ кодирования аминокислотной последовательности белков при помощи последовательности нуклеотидов.
Свойства генетического кода: 1. универсальность (принцип записи един для всех живых организмов) 2. триплетность (считываются три, рядом расположенные нуклеотида) 3. специфичность (1 триплет соответствует ТОЛЬКО ОДНОЙ аминокислоте) 4. вырожденность (избыточность) (1 аминокислота может кодироваться несколькими триплетами) 5. неперекрываемость (считывание происходит триплет за триплетом без "пробелов" и областей перекрывания, т.е. 1 нуклеотид НЕ может входить в состав двух триплетов).
Вопрос 13 (Характеристика этапов биосинтеза белка у про- и эукариот):
Биосинтез белка у эукариот
Транскрипция,постранскрипция, трансляция и посттрансляция. 1.Транскрипция заключается в создании "копии одного гена" - молекулы пре-и-РНК (пре-м-РНК).Происходит разрыв водородных связей между азотистыми основаниями, присоединения к гену-промотору РНК полимеразы, которая "подбирает" нуклеотиды по принципу комплементарности, и антипараллельности. Гены у эукариот содержат участки, содержащие информацию, - экзоны и неинформативные участки - экзоны. В результате транскрипции создается "копия" гена, которая содержит как экзоны, так и интроны. Поэтому молекула, синтезирующаяся в результате транскрипции у эукариот - незрелая и-РНК (пре-и-РНК). 2.Период посттранскрипции он называется процессинг, который заключается в созревании и-РНК. Происходит: Вырезание интронов и сшивание (сплайсинг) экзонов (сплайсинг называется альтернативным, если экзоны соединяются в другой последовательности, чем были изначально в молекуле ДНК). Происходит "модификация концов" пре-и-РНК: на начальном участке - лидере (5") образуется колпачок или кэп - для узнавания и связывания с рибосомой, на конце 3" - трейлере образуется polyА (множество адениловых оснований) - для транспорта и-РНК из мембраны ядра в цитоплазму. Это зрелая м РНК.
3. Трансляция: -Инициация -связывание и-РНК с малой субъединицей рибосомы -попадание стартового триплета и-РНК - АУГ в аминоацильный центр рибосомы -объединение 2-ух субъединиц рибосомы (большой и малой). -Элонгация АУГ попадает в пептидильный центр, а в аминоацильный центр попадает второй триплет, потом две тРНК с определенными аминокислотами поступают в оба центра рибосомы. В случае комплементарности триплетов на и-РНК (кодона) и т-РНК (антикодон, на центральной петле молекулы т-РНК) между ними образуются водородные связи и данные т-РНК с соответствующими АМК "фиксируются" в рибосоме. Между АМК, прикрепленными к двум т-РНК, возникает пептидная связь, а связь между первой АМК и первой т-РНК разрушается. Рибосмома делает "шаг" по и-РНК ("передвигается на один триплет). Таким образом, вторая т-РНК, к которой прикреплены уже две АМК, перемещается в пептидильный центр, а в аминоацильном центре оказывается третий триплет и-РНК, куда из цитоплазмы поступает следующая т-РНК с соответствующей АМК. Процесс повторяется... до тех пор, пока в аминоацильный центр не попадет один из трех стоп-кодонов (УАА, УАГ, УГА), которые не соответствуют ни одной аминокислоте
Терминация - окончание сборки полипептидной цепи. Результат трансляции - образование полипептидной цепи, т.е. первичной структуры белка. 4. Посттрансляция приобретение молекулой белка соответствующей конформации - вторичной, третичной, четвертичной структур. Особенности биосинтеза белка у прокариот: а) все этапы биосинтеза происходят в цитоплазме, б) отсутствие экзон-интронной организации генов, вследствие чего в результате транскрипции образуется зрелая полицистронная м-РНК, в) транскрипция сопряжена с трансляцией, г) имеется только 1 вид РНК-полимеразы (единый РНК-полимеразный комплекс), тогда как у эукариот 3 вида РНК-полимераз, осуществляющих транскрипцию разных видов РНК.
Все живые организмы, за исключением вирусов, состоят из клеток. При этом вирусы нельзя назвать в полной мере самостоятельными живыми организмами. Для размножения им нужны клетки, т. е. они заражают другие организмы. Таким образом, мы можем сказать, что жизнь в полной мере может осуществляться только в клетках.
Клетки разных живых организмов имеют общий план строения, многие процессы в них протекают одинаково. Однако между клетками организмов, принадлежащих к разным царствам , есть некоторые ключевые различия. Так, например, клетки бактерий не имеют ядер. У клеток животных и растений ядра есть. Но у них есть другие различия.
У клеток растений в отличие от животных есть три выраженных особенности. Это наличие клеточной стенки , пластид и центральной вакуоли.
И клетки растений, и клетки животных окружены клеточной мембраной. Она ограничивает содержимое клетки от внешней среды, пропускает одни вещества и не пропускает другие. При этом у растений с внешней стороны от мембраны есть еще клеточная стенка , или клеточная оболочка . Она достаточно жесткая и придает растительной клетке форму. Благодаря клеточным стенкам растениям не нужен скелет. Без них растения бы наверно «растеклись» по земле. А так даже трава может стоять вертикально. Чтобы вещества могли проникать через клеточную оболочку, в ней есть поры. Также через эти поры клетки контактируют между собой, образуя цитоплазматические мостики. Клеточная стенка состоит из целлюлозы.
Пластиды есть только у клеток растений. К пластидам относятся хлоропласты, хромопласты и лейкопласты. Наиболее важное значение имеют хлоропласты . В них протекает процесс фотосинтеза, при котором из неорганических веществ синтезируются органические. Животные синтезировать органические вещества из неорганических не могут. Они получают с пищей готовые органические вещества, при необходимости расщепляют их до более простых и синтезируют уже свои органические вещества. Несмотря на то, что растения могут фотосинтезировать, подавляющее большинство органических веществ в них образуется также из других органических. Однако родоначальником всего органического в них служит органическое вещество, которое получается в хлоропластах из неорганических веществ. Это вещество - глюкоза.
Крупная центральная вакуоль характерна только для растительных клеток. В животных клетках тоже бывают вакуоли. Однако по мере роста клетки они не сливаются в одну большую вакуоль, которая оттесняет все остальное содержимое клетки к мембране. Именно так происходит у растений. Вакуоль содержит клеточный сок, содержащий в основном запасные вещества. Крупная вакуоль создает внутреннее давление на клеточную мембрану. Таким образом наряду с клеточной оболочкой она поддерживает форму клетки.
Запасным питательным веществом углеводного типа в растительных клетках является крахмал, а в животных - гликоген. Крахмал и гликоген очень похожи по строению.
У животных клеток также есть «свои» органеллы, которых нет у высших растений. Это центриоли. Они участвуют в процессе деления клеток.
Остальные органеллы у растительных и животных клеток сходны по строению и функциям. Это митохондрии, комплекс Гольджи, ядро, эндоплазматическая сеть, рибосомы и некоторые другие.
На заре развития жизни на Земле все клеточные формы были представлены бактериями. Они всасывали органические вещества, растворённые в первичном океане, через поверхность тела.
Со временем некоторые бактерии приспособились производить органические вещества из неорганических. Для этого они использовали энергию солнечного света. Возникла первая экологическая система, в которой эти организмы были производителями. В результате этого в атмосфере Земли появился кислород, выделяемый этими организмами. С его помощью можно из той же самой пищи получить гораздо больше энергии, а добавочную энергию использовать на усложнение строения тела: разделение тела на части.
Одно из важных достижений жизни — разделение ядра и цитоплазмы. В ядре находится наследственная информация. Специальная мембрана вокруг ядра позволила защитить от случайных повреждений. По мере необходимости цитоплазма получает из ядра команды, направляющие жизнедеятельность и развитие клетки.
Организмы, у которых ядро отделено от цитоплазмы, образовали надцарство ядерных (к ним относятся — растения, грибы, животные).
Таким образом, клетка — основа организации растений и животных — возникла и развилась в ходе биологической эволюции.
Даже не вооружённым глазом, а ещё лучше под лупой можно видеть, что мякоть зрелого арбуза состоит из очень мелких крупинок, или зёрнышек. Это клетки — мельчайшие «кирпичики», из которых состоят тела всех живых организмов, в том числе и растительных.
Жизнь растения осуществляется соединённой деятельностью его клеток, создающих единое целое. При многоклеточности частей растения существует физиологическое разграничение их функций, специализация различных клеток в зависимости от местоположения их в теле растения.
Растительная клетка отличается от животной тем, что имеет плотную оболочку, покрывающую внутреннее содержимое со всех сторон. Клетка не является плоской (как её принято изображать), она скорей всего похожа на очень маленький пузырёк, наполненный слизистым содержимым.
Строение и функции растительной клетки
Рассмотрим клетку как структурно-функциональную единицу организма. Снаружи клетка покрыта плотной клеточной стенкой, в которой имеются более тонкие участки — поры. Под ней находится очень тонкая плёнка — мембрана, покрывающая содержимое клетки — цитоплазму. В цитоплазме есть полости — вакуоли, заполненные клеточным соком. В центре клетки или около клеточной стенки расположено плотное тельце — ядро с ядрышком. От цитоплазмы ядро отделено ядерной оболочкой. По всей цитоплазме распределены мелкие тельца — пластиды.
Строение растительной клетки
Строение и функции органоидов растительной клетки
| Органоид | Рисунок | Описание | Функция | Особенности |
Клеточная стенка или плазматическая мембрана | Бесцветная, прозрачная и очень прочная | Пропускает в клетку и выпускает из клетки вещества. | Клеточная мембрана полупроницаемая |
|
Цитоплазма | Густое тягучее вещество | В ней располагаются все другие части клетки | Находится в постоянном движении |
|
Ядро (важная часть клетки) | Округлое или овальное | Обеспечивает передачу наследственных свойств дочерним клеткам при делении | Центральная часть клетки |
|
Сферической или неправильной формы | Принимает участие в синтезе белка | |||
 | Резервуар, отделённый от цитоплазмы мембраной. Содержит клеточный сок | Накапливаются запасные питательные вещества и продукты жизнедеятельности ненужные клетке. | По мере роста клетки мелкие вакуоли сливаются в одну большую (центральную) вакуоль |
|
Пластиды | Хлоропласты | Используют световую энергию солнца и создают органические из неорганических | Форма дисков, отграниченных от цитоплазмы двойной мембраной |
|
Хромопласты | Образуются в результате накопления каротиноидов | Жёлтые, оранжевые или бурые |
||
 | Лейкопласты | Бесцветные пластиды | ||
Ядерная оболочка | Состоит из двух мембран (наружная и внутренняя) с порами | Отграничивает ядро от цитоплазмы | Даёт возможность осуществляться обмену между ядром и цитоплазмой |
Живая часть клетки — это ограниченная мембраной, упорядоченная, структурированная система биополимеров и внутренних мембранных структур, участвующих в совокупности метаболических и энергетических процессов, осуществляющих поддержание и воспроизведение всей системы в целом.
Важной особенностью является то, что в клетке нет открытых мембран со свободными концами. Клеточные мембраны всегда ограничивают полости или участки, закрывая их со всех сторон.
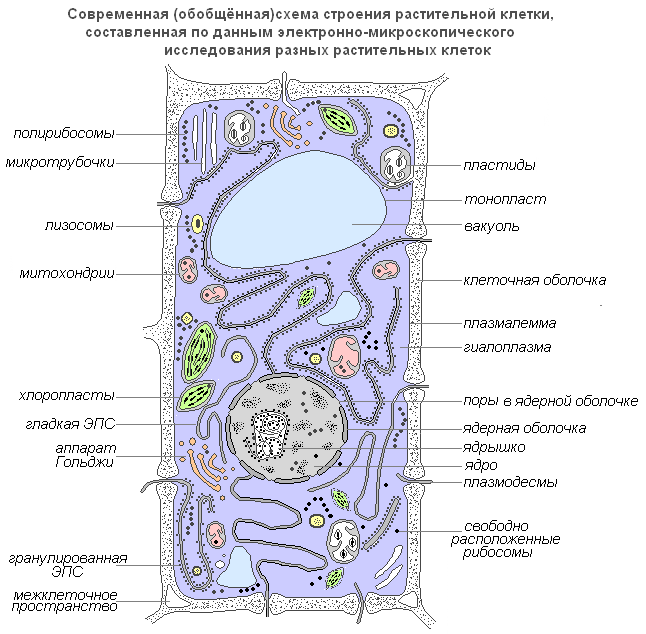
Современная обобщенная схема растительной клетки
Плазмалемма (наружная клеточная мембрана) — ультрамикроскопическая плёнка толщиной 7,5 нм., состоящая из белков, фосфолипидов и воды. Это очень эластичная плёнка, хорошо смачивающаяся водой и быстро восстанавливающая целостность после повреждения. Имеет универсальное строение, т.е.типичное для всех биологических мембран. У растительных клеток снаружи от клеточной мембраны находится прочная, создающая внешнюю опору и поддерживающая форму клетки клеточная стенка. Она состоит из клетчатки (целлюлозы) — нерастворимого в воде полисахарида.
Плазмодесмы растительной клетки, представляют собой субмикроскопические канальцы, пронизывающие оболочки и выстланные плазматической мембраной, которая таким образом переходит из одной клетки в другую, не прерываясь. С их помощью происходит межклеточная циркуляция растворов, содержащих органические питательные вещества. По ним же идёт передача биопотенциалов и другой информации.
Порами называют отверстия во вторичной оболочке, где клетки разделяют лишь первичная оболочка и срединная пластинка. Участки первичной оболочки и срединную пластинку, разделяющие соседствующие поры смежных клеток, называют поровой мембраной или замыкающей пленкой поры. Замыкающую пленку поры пронизывают плазмодесменные канальцы, но сквозного отверстия в порах обычно не образуется. Поры облегчают транспорт воды и растворенных веществ от клетки к клетке. В стенках соседних клеток, как правило, одна против другой, образуются поры.
Клеточная оболочка имеет хорошо выраженную, относительно толстую оболочку полисахаридной природы. Оболочка растительной клетки продукт деятельности цитоплазмы. В её образовании активное участие принимает аппарат Гольджи и эндоплазматическая сеть.
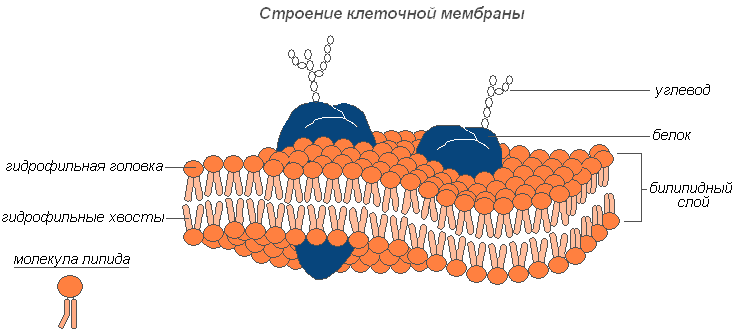
Строение клеточной мембраны
Основу цитоплазмы составляет ее матрикс, или гиалоплазма, — сложная бесцветная, оптически прозрачная коллоидная система, способная к обратимым переходам из золя в гель. Важнейшая роль гиалоплазмы заключается в объединении всех клеточных структур в единую систему и обеспечении взаимодействия между ними в процессах клеточного метаболизма.
Гиалоплазма (или матрикс цитоплазмы) составляет внутреннюю среду клетки. Состоит из воды и различных биополимеров (белков, нуклеиновых кислот, полисахаридов, липидов), из которых основную часть составляют белки различной химической и функциональной специфичности. В гиалоплазме содержатся также аминокислоты, моносахара, нуклеотиды и другие низкомолекулярные вещества.
Биополимеры образуют с водой коллоидную среду, которая в зависимости от условий может быть плотной (в форме геля) или более жидкой (в форме золя), как во всей цитоплазме, так и в отдельных ее участках. В гиалоплазме локализуются и взаимодействуют между собой и средой гиалоплазмы различные органеллы и включения. При этом расположение их чаще всего специфично для определенных типов клеток. Через билипидную мембрану гиалоплазма взаимодействует с внеклеточной средой. Следовательно, гиалоплазма является динамической средой и играет важную роль в функционировании отдельных органелл и жизнедеятельности клеток в целом.
Цитоплазматические образования – органеллы
Органеллы (органоиды) — структурные компоненты цитоплазмы. Они имеют определённую форму и размеры, являются обязательными цитоплазматическими структурами клетки. При их отсутствии или повреждении клетка обычно теряет способность к дальнейшему существованию. Многие из органоидов способны к делению и самовоспроизведению. Размеры их настолько малы, что их можно видеть только в электронный микроскоп.
Ядро
Ядро — самая заметная и обычно самая крупная органелла клетки. Оно впервые было подробно исследовано Робертом Броуном в 1831 году. Ядро обеспечивает важнейшие метаболические и генетические функции клетки. По форме оно достаточно изменчиво: может быть шаровидным, овальным, лопастным, линзовидным.
Ядро играет значительную роль в жизни клетки. Клетка, из которой удалили ядро, не выделяет более оболочку, перестаёт расти и синтезировать вещества. В ней усиливаются продукты распада и разрушения, вследствие этого она быстро погибает. Образование нового ядра из цитоплазмы не происходит. Новые ядра образуются только делением или дроблением старого.
Внутреннее содержимое ядра составляет кариолимфа (ядерный сок), заполняющая пространство между структурами ядра. В нём находится одно или несколько ядрышек, а также значительное количество молекул ДНК, соединённых со специфическими белками — гистонами.

Строение ядра
Ядрышко
Ядрышко — как и цитоплазма, содержит преимущественно РНК и специфические белки. Важнейшая его функция заключается в том, что в нём происходит формирование рибосом, которые осуществляют синтез белков в клетке.
Аппарат Гольджи
Аппарат Гольджи — органоид, имеющий универсальное распространение во всех разновидностях эукариотических клеток. Представляет собой многоярусную систему плоских мембранных мешочков, которые по периферии утолщаются и образуют пузырчатые отростки. Он чаще всего расположен вблизи ядра.

Аппарат Гольджи
В состав аппарата Гольджи обязательно входит система мелких пузырьков (везикул), которые отшнуровываются от утолщённых цистерн (диски) и располагаются по периферии этой структуры. Эти пузырьки играют роль внутриклеточной транспортной системы специфических секторных гранул, могут служить источником клеточных лизосом.
Функции аппарата Гольджи состоят также в накоплении, сепарации и выделении за пределы клетки с помощью пузырьков продуктов внутриклеточного синтеза, продуктов распада, токсических веществ. Продукты синтетической деятельности клетки, а также различные вещества, поступающие в клетку из окружающей среды по каналам эндоплазматической сети, транспортируются к аппарату Гольджи, накапливаются в этом органоиде, а затем в виде капелек или зёрен поступают в цитоплазму и либо используются самой клеткой, либо выводятся наружу. В растительных клетках Аппарат Гольджи содержит ферменты синтеза полисахаридов и сам полисахаридный материал, который используется для построения клеточной оболочки. Предполагают, что он участвует в образовании вакуолей. Аппарат Гольджи был назван так в честь итальянского учёного Камилло Гольджи, впервые обнаружившего его в 1897 году.
Лизосомы
Лизосомы представляют собой мелкие пузырьки, ограниченные мембраной основная функция которых — осуществление внутриклеточного пищеварения. Использование лизосомного аппарата происходит при прорастании семени растения (гидролиз запасных питательных веществ).
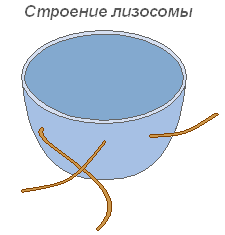
Строение лизосомы
Микротрубочки
Микротрубочки — мембранные, надмолекулярные структуры, состоящие из белковых глобул, расположенных спиральными или прямолинейными рядами. Микротрубочки выполняют преимущественно механическую (двигательную) функцию, обеспечивая подвижность и сокращаемость органоидов клетки. Располагаясь в цитоплазме, они придают клетке определённую форму и обеспечивают стабильность пространственного расположения органоидов. Микротрубочки способствуют перемещению органоидов в места, которые определяются физиологическими потребностями клетки. Значительное количество этих структур расположено в плазмалемме, вблизи клеточной оболочки, где они участвуют в формировании и ориентации целлюлозных микрофибрилл оболочек растительных клеток.

Строение микротрубочки
Вакуоль
Вакуоль — важнейшая составная часть растительных клеток. Она представляет собой своеобразную полость (резервуар) в массе цитоплазмы, заполненную водным раствором минеральных солей, аминокислот, органических кислот, пигментов, углеводов и отделённую от цитоплазмы вакуолярной мембраной — тонопластом.
Цитоплазма заполняет всю внутреннюю полость только у самых молодых растительных клеток. С ростом клетки существенно изменяется пространственное расположение вначале сплошной массы цитоплазмы: у неё появляются заполненные клеточным соком небольшие вакуоли, и вся масса становится ноздреватой. При дальнейшем росте клетки отдельные вакуоли сливаются, оттесняя к периферии прослойки цитоплазмы, в результате чего в сформированной клетке находится обычно одна большая вакуоль, а цитоплазма со всеми органеллами располагаются около оболочки.
Водорастворимые органические и минеральные соединения вакуолей обусловливают соответствующие осмотические свойства живых клеток. Этот раствор определённой концентрации является своеобразным осмотическим насосом для регулируемого проникновения в клетку и выделения из неё воды, ионов и молекул метаболитов.
В комплексе со слоем цитоплазмы и её мембранами, характеризующимися свойствами полупроницаемости, вакуоль образует эффективную осмотическую систему. Осмотически обусловленными являются такие показатели живых растительных клеток, как осмотический потенциал, сосущая сила и тургорное давление.

Строение вакуоли
Пластиды
Пластиды — самые крупные (после ядра) цитоплазматические органоиды, присущие только клеткам растительных организмов. Они не найдены только у грибов. Пластиды играют важную роль в обмене веществ. Они отделены от цитоплазмы двойной мембранной оболочкой, а некоторые их типы имеют хорошо развитую и упорядоченную систему внутренних мембран. Все пластиды едины по происхождению.
Хлоропласты — наиболее распространённые и наиболее функционально важные пластиды фотоавтотрофных организмов, которые осуществляют фотосинтетические процессы, приводящие в конечном итоге к образованию органических веществ и выделению свободного кислорода. Хлоропласты высших растений имеют сложное внутреннее строение.

Строение хлоропласта
Размеры хлоропластов у разных растений неодинаковы, но в среднем диаметр их составляет 4-6 мкм. Хлоропласты способны передвигаться под влиянием движения цитоплазмы. Кроме того, под воздействием освещения наблюдается активное передвижение хлоропластов амебовидного типа к источнику света.
Хлорофилл — основное вещество хлоропластов. Благодаря хлорофиллу зелёные растения способны использовать световую энергию.
Лейкопласты (бесцветные пластиды) представляют собой чётко обозначенные тельца цитоплазмы. Размеры их несколько меньше, чем размеры хлоропластов. Более и однообразна и их форма, приближающая к сферической.
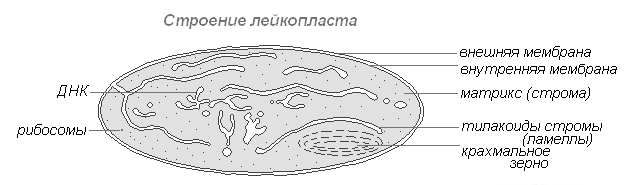
Строение лейкопласта
Встречаются в клетках эпидермиса, клубнях, корневищах. При освещении очень быстро превращаются в хлоропласты с соответствующим изменением внутренней структуры. Лейкопласты содержат ферменты, с помощью которых из излишков глюкозы, образованной в процессе фотосинтеза, в них синтезируется крахмал, основная масса которого откладывается в запасающих тканях или органах (клубнях, корневищах, семенах) в виде крахмальных зёрен. У некоторых растений в лейкопластах откладываются жиры. Резервная функция лейкопластов изредка проявляется в образовании запасных белков в форме кристаллов или аморфных включений.
Хромопласты в большинстве случаев являются производными хлоропластов, изредка — лейкопластов.

Строение хромопласта
Созревание плодов шиповника, перца, помидоров сопровождается превращением хлоро- или лейкопластов клеток мякоти в каратиноидопласты. Последние содержат преимущественно жёлтые пластидные пигменты — каратиноиды, которые при созревании интенсивно синтезируются в них, образуя окрашенные липидные капли, твёрдые глобулы или кристаллы. Хлорофилл при этом разрушается.
Митохондрии
Митохондрии — органеллы, характерные для большинства клеток растений. Имеют изменчивую форму палочек, зёрнышек, нитей. Открыты в 1894 году Р. Альтманом с помощью светового микроскопа, а внутреннее строение было изучено позднее с помощью электронного.

Строение митохондрии
Митохондрии имеют двухмембранное строение. Внешняя мембрана гладкая, внутренняя образует различной формы выросты — трубочки в растительных клетках. Пространство внутри митохондрии заполнено полужидким содержимым (матриксом), куда входят ферменты, белки, липиды, соли кальция и магния, витамины, а также РНК, ДНК и рибосомы. Ферментативный комплекс митохондрий ускоряет работу сложного и взаимосвязанного механизма биохимических реакций, в результате которых образуется АТФ. В этих органеллах осуществляется обеспечение клеток энергией — преобразование энергии химических связей питательных веществ в макроэргиеские связи АТФ в процессе клеточного дыхания. Именно в митохондриях происходит ферментативное расщепление углеводов, жирных кислот, аминокислот с освобождением энергии и последующим превращением её в энергию АТФ. Накопленная энергия расходуется на ростовые процессы, на новые синтезы и т. д. Митохондрии размножаются делением и живут около 10 дней, после чего подвергаются разрушению.
Эндоплазматическая сеть
Эндоплазматическая сеть — сеть каналов, трубочек, пузырьков, цистерн, расположенных внутри цитоплазмы. Открыта в 1945 году английским учёным К. Портером, представляет собой систему мембран, имеющих ультрамикроскопическое строение.

Строение эндоплазматической сети
Вся сеть объединена в единое целое с наружной клеточной мембраной ядерной оболочки. Различают ЭПС гладкую и шероховатую, несущую на себе рибосомы. На мембранах гладкой ЭПС находятся ферментные системы, участвующие в жировом и углеводном обмене. Этот тип мембран преобладает в клетках семян, богатых запасными веществами (белками, углеводами, маслами), рибосомы прикрепляются к мембране гранулярной ЭПС, и во время синтеза белковой молекулы полипептидная цепочка с рибосомами погружается в канал ЭПС. Функции эндоплазматической сети очень разнообразны: транспорт веществ как внутри клетки, так и между соседними клетками; разделение клетки на отдельные секции, в которых одновременно проходят различные физиологические процессы и химические реакции.
Рибосомы
Рибосомы — немембранные клеточные органоиды. Каждая рибосома состоит из двух не одинаковых по размеру частичек и может делиться на два фрагмента, которые продолжают сохранять способность синтезировать белок после объединения в целую рибосому.

Строение рибосомы
Рибосомы синтезируются в ядре, затем покидают его, переходя в цитоплазму, где прикрепляются к наружной поверхности мембран эндоплазматической сети или располагаются свободно. В зависимости от типа синтезируемого белка рибосомы могут функционировать по одиночке или объединяться в комплексы — полирибосомы.
2. Основные химические компоненты протопласта . Органические вещества клетки. Белки - биополимеры , образованные аминокислотами , составляют 40-50% сухой массы протопласта. Они участвуют в построении структуры и функциях всех органелл . В химическом отношении белки подразделяются на простые (протеины) и сложные (протеиды). Сложные белки могут образовывать комплексы с липидами - липопротеиды, с углеводами - гликопротеиды, с нуклеиновыми кислотами - нуклеопротеиды и т.д.
Белки входят в состав ферментов (энзимов), регулирующих все жизненно важные процессы.
Цитоплазма представляет собой густой прозрачный коллоидный раствор. В зависимости от выполняемых физиологических функций каждая клетка имеет свой химический состав. Основу цитоплазмы составляет ее гиалоплазма, или матрикс, роль которой заключается в объединении всех клеточных структур в единую систему и обеспечении взаимодействия между ними. Цитоплазма имеет щелочную реакцию среды и на 60-90 % состоит из воды, в которой растворены различные вещества: до 10-20 % белков, 2-3 % жироподобных веществ, 1,5 % органических и 2-3 % неорганических соединений. В цитоплазме осуществляется важнейший физиологический процесс - дыхание, или гликолиз, в результате которого происходит расщепление глюкозы без доступа кислорода в присутствии ферментов с освобождением энергии и образованием воды и диоксида углерода.
Цитоплазма пронизана мембранами - тончайшими пленками фосфолипидного строения. Мембраны образуют эндоплазматическую сеть - систему мелких канальцев и полостей, образующих сеть. Эндоплазматическая сеть называется шероховатой (гранулярной), если на мембранах канальцев и полостей находятся рибосомы или группы рибосом, которые выполняют синтез белка. Если эндоплазматическая сеть лишена рибосом, то называется гладкой (агранулярной). На мембранах гладкой эндоплазматической сети синтезируются липиды и углеводы.
Аппарат Гольджи - система уплощенных цистерн, лежащих параллельно и ограниченных двойными мембранами. От концов цистерн отшнуровываются пузырьки, через которые удаляются конечные или ядовитые продукты жизнедеятельности клетки, обратно же в диктиосомы поступают вещества, необходимые для синтеза сложных углеводов (полисахаридов) на построение клеточной стенки. Также комплекс Гольджи участвует в формировании вакуолей. Одно из важнейших биологических свойств цитоплазмы - циклоз (способность к движению), интенсивность которого зависит от температуры, степени освещения, снабжения кислородом и других факторов.
Рибосомы - мельчайшие частицы (от 17 до 23 нм), образованные рибонуклеопротеидами и молекулами белка. Они присутствуют в цитоплазме, ядре, митохондриях, пластидах; бывают одиночными и групповыми (полисомы). Рибосомы - центры синтеза белка.
Митохондрии - «энергетические станции» всех эукариотических клеток. Форма их разнообразна: от округлых до цилиндрических и даже палочковидных телец. Численность их - от нескольких десятков до нескольких тысяч в каждой клетке. Размеры не более 1 мкм. Снаружи митохондрии окружены двухмембранной оболочкой. Внутренняя мембрана представлена в виде пластинчатых выростов - крист. Размножаются путем деления.
Основная функция митохондрий - участие в дыхании клетки с помощью ферментов. В митохондриях в результате реакции окислительного фосфорилирования синтезируются богатые энергией молекулы аденозинтрифосфорной кислоты (АТФ). Механизм окислительного фосфорилирования был открыт английским биохимиком П. Митчелом в 1960 г.
Пластиды. Эти органеллы, характерные только для растений, встречаются во всех живых растительных клетках. Пластиды - относительно крупные (4-10 мкм) живые растительные тельца разной формы и окраски. Различают три типа пластид: 1) хлоропласты, окрашенные в зеленый цвет; 2) хромопласты, окрашенные в желто-красные цвета; 3) лейкопласты, не имеющие окраски.
Хлоропласты встречаются во всех зеленых органах растений. У высших растений пластид в клетках несколько десятков, у низших (водорослей) - 1-5. Они крупные, разнообразны по форме. В хлоропластах содержится до 75 % воды, белки, липиды, нуклеиновые кислоты, ферменты и красящие вещества - пигменты. Для образования хлорофилла необходимы определенные условия - свет, соли железа и магния в почве. От цитоплазмы хлоропласт отделен двойной мембранной оболочкой; тело его состоит из бесцветной мелкозернистой стромы. Строма пронизана параллельно расположенными пластинками - ламеллами, дисками. Диски собраны в стопки - граны. Основная функция хлоропластов - фотосинтез.
Хромопласты встречаются в корнеплодах моркови, плодах многих растений (облепиха, шиповник, рябина и др.), в зеленых листьях шпината, крапивы, в цветках (розы, гладиолусы, календула), окраска которых зависит от присутствия в них пигментов каротиноидов: каротина - оранжево-красного цвета и ксантофилла - желтого цвета.
Лейкопласты - бесцветные пластиды, пигменты отсутствуют. Они представляют собой белковые вещества в виде шаровидных, веретонообразных зернышек, концентрирующихся вокруг ядра. В них осуществляется синтез и накопление запасных питательных веществ, в основном крахмала, белков и жиров. Лейкопласты находятся в цитоплазме, эпидерме, молодых волосках, подземных органах растений и в тканях зародыша семени.
Пластиды могут переходить из одного вида в другой.
Ядро.
Ядро - одно из главных органелл эукариотической клетки. В растительной клетке одно ядро. В ядре хранится и воспроизводится наследственная информация. Размеры ядра у разных растений разные, от 2-3 до 500 мкм. Форма чаще округлая или чечевицеобразная. В молодых клетках ядро крупнее, чем в старых, и занимает центральное положение. Ядро окружено двойной мембраной с порами, регулирующими обмен веществ. Наружная мембрана объединена с эндоплазматической сетью. Внутри ядра заключен ядерный сок - кариоплазма с хроматином, ядрышками и рибосомами. Хроматин - бесструктурная среда из особых нуклеопротеидных нитей, богатых ферментами.
В хроматине сосредоточена основная масса ДНК. В процессе клеточного деления хроматин превращается в хромосомы - носители генов. Хромосомы образованы двумя одинаковыми нитями ДНК - хроматидами. Каждая хромосома в середине имеет перетяжку - центромеру. Число хромосом у разных растений неодинакова: от двух до нескольких сотен. Каждый вид растений имеет постоянный набор хромосом. В хромосомах синтезируются нуклеиновые кислоты, необходимые для образования белков. Совокупность количественных и качественных признаков хромосомного набора клетки называют кариотипом. Изменение числа хромосом происходит в результате мутаций. Наследственное кратное увеличение числа хромосом у растений получило название полиплоидии.
Ядрышки - сферические, довольно плотные тельца диаметром 1-3 мкм. В ядре содержатся 1-2, иногда несколько ядрышек. Ядрышко является основным носителем РНК ядра. Основная функция ядрышка - синтез рРНК.
Деление ядра и клетки. Размножение клеток происходит путем их деления. Период между двумя последовательными делениями составляет клеточный цикл. При делении клеток наблюдается рост растения и увеличение его общей массы. Существуют три способа деления клеток: митоз, или кариокинез (непрямое деление), мейоз (редукционное деление) и амитоз (прямое деление).
Митоз характерен для всех клеток органов растений, кроме половых. В результате митоза растет и увеличивается общая масса растения. Биологическое значение митоза заключается в строго одинаковом распределении редуплицированных хромосом между дочерними клетками, что обеспечивает образование генетически равноценных клеток. Митоз впервые был описан русским ботаником И.Д.Чистяковым в 1874 г. В процессе митоза выделяют несколько фаз: профазу, метафазу, анафазу и телофазу. Промежуток между двумя делениями клетки называется интерфазой. В интерфазе осуществляется общий рост клетки, редупликация органоидов, синтез ДНК, формирование и подготовка структур к началу митотического деления.
Профаза - самая длительная фаза митоза. В профазе хромосомы становятся видны в световой микроскоп. В профазе ядро претерпевает два изменения: 1. стадия плотного клубка; 2. стадия рыхлого клубка. В стадии плотного клубка хромосомы становятся видны в световой микроскоп, раскручиваются из клубка или из спирали и вытягиваются. Каждая хромосома состоит из двух хроматид, расположенных параллельно друг другу. Постепенно они укорачиваются, утолщаются и обособляются, исчезают ядерная оболочка и ядрышко. Ядро увеличивается в объеме. На противоположных полюсах клетки образуется ахроматиновое веретено - веретено деления, состоящее из неокрашивающихся нитей, протягивающихся от полюсов клетки (стадия рыхлого клубка).
В метафазе заканчивается формирование веретена деления, хромосомы приобретают определенную форму того или иного вида растения и собираются в одной плоскости - экваториальной, на месте бывшего ядра. Ахроматиновое веретено постепенно сокращается, и хроматиды начинают отделяться друг от друга, оставаясь связанными в области центромеры.
В анафазе происходит деление центромеры. Образовавшиеся сестринские центромеры и хроматиды направляются к противоположным полюсам клетки. Самостоятельные хроматиды становятся дочерними хромосомами, и, следовательно, их будет точно столько, сколько в материнской клетке.
Телофаза - последняя фаза деления клетки, когда дочерние хромосомы достигают полюсов клетки, постепенно исчезает веретено деления, хромосомы удлиняются и становятся плохо заметными в световой микроскоп, в экваториальной плоскости формируется срединная пластинка. Постепенно образуется клеточная стенка и одновременно - ядрышки и ядерная оболочка вокруг двух новых ядер (1. стадия рыхлого клубка; 2. стадия плотного клубка). Образовавшиеся клетки вступают в очередную интерфазу.
Длительность митоза примерно 1-2 часа. Процесс от момента образования срединной пластинки до формирования новой клетки называют цитокинезом. Дочерние клетки в два раза мельче материнских, но затем они растут и достигают размеров материнской клетки.
Мейоз . Впервые был открыт русским ботаником В.И. Беляевым в 1885 г. Этот тип деления клеток связан с образованием спор и гамет, или половых клеток, имеющих гаплоидное число хромосом (n). Сущность его заключается в уменьшении (редукции) числа хромосом в 2 раза в каждой образовавшейся после деления клетке. Мейоз состоит из двух следующих друг за другом делений. Мейоз в отличие от митоза состоит из двух видов деления: редукционного (увеличение); экватоционного (митотическое деление). Редукционное деление происходит при первом делении, которое состоит из нескольких фаз: профаза I, метафаза I, анафаза I, телофаза I. В экватоционном делении различают: профаза II, метафаза II, анафаза II, телофаза II. В редукционном делении существует интерфаза.
Профаза I. Хромосомы имеют форму длинных двойных нитей. Хромосома состоит из двух хроматид. Это стадия лептонемы. Затем гомологичные хромосомы притягиваются друг к другу, образуя пары - биваленты. Эта стадия называется зигонемой. Спаренные гомологичные хромосомы состоят из четырех хроматид, или тетрад. Хроматиды могут быть расположены параллельно друг другу либо перекрещиваться между собой, обмениваясь участками хромосом. Эта стадия получила название кроссинговера. В следующей стадии профазы I - пахинеме, хромосомные нити утолщаются. В следующей стадии - диплонеме - тетрады хроматид укорачиваются. Конъюгирующие хромосомы сближаются друг с другом так, что становятся неразличимыми. Исчезают ядрышко и ядерная оболочка, формируется ахроматиновове веретено. В последней стадии - диакинезе - биваленты направляются к экваториальной плоскости.
Метафаза I. Биваленты располагаются по экватору клетки. Каждая хромосома прикреплена ахроматиновым веретеном к центромере.
Анафаза I. Происходит сокращение нитей ахроматинового веретена, и гомологичные хромосомы в каждом биваленте расходятся к противоположным полюсам, причем на каждом полюсе окажется половинное число хромосом материнской клетки, т.е. происходит уменьшение (редукция) числа хромосом и образуются два гаплоидных ядра.
Телофаза I. Эта фаза слабо выражена. Хромосомы деконденсируются; ядро принимает вид интерфазного, но в нем не происходит удвоения хромосом. Эта стадия называется интеркинезом. Она непродолжительная, у некоторых видов отсутствует, и тогда клетки сразу после телофазы I переходят в профазу II.
Второе мейотическое деление происходит по типу митоза.
Профаза II. Наступает быстро, вслед за телофазой I. Видимых изменений в ядре не происходит и сущность этой стадии заключается в том, что происходит рассасывание ядерных оболочек и появление четырех полюсов деления. Возле каждого ядра возникает два полюса.
Метафаза II. Удвоенные хромосомы выстраиваются у своих экваторов и стадия носит название стадии материнской звезды или экваториальной пластинки. От каждого полюса деления отходят нити веретена деления, которые прикрепляются к хроматидам.
Анафаза II. Полюса делений натягивают нити веретена деления, которые начинают рассасываться и натягивать удвоенные хромосомы. Наступает момент разрыва хромосом и расхождения их к четырем полюсам.
Телофаза II. Вокруг каждого полюса у хромосом происходит стадия рыхлого клубка и стадия плотного клубка. После чего рассасываются центриоли и вокруг хромосом восстанавливаются ядерные оболочки и ядрышки. После чего делится и цитоплазма.
Итогом мейоза является образование четырех дочерних клеток из одной материнской с гаплоидным набором хромосом.
Для каждого вида растений характерно постоянное число хромосом и постоянная их форма. Среди высших растений часто встречается явление полиплоидии, т.е. многократное повторение в ядре одного набора хромосом (триплоиды, тетераплоиды и т.д.).
В старых и больных клетках растений можно наблюдать прямое (амитоз) деление ядра путем простой его перетяжки на две части с произвольным количеством ядерного вещества. Впервые это деление было описано Н. Железновым в 1840 г.
Производные протопласта.
К производным протопласта относятся:
1) вакуоли;
2) включения;
3) клеточная стенка;
4) физиологически активные вещества: ферменты, витамины, фитогормоны и др.;
5) продукты обмена веществ.
Вакуоли - полости в протопласте - производные эндоплазматической сети. Они ограничены мембраной - тонопластом и заполнены клеточным соком. Клеточный сок накапливается в каналах эндоплазматической сети в виде капелек, которые затем сливаются, образуя вакуоли. В молодых клетках содержится много мелких вакуолей, в старой клетке обычно присутствует одна крупная вакуоль. В клеточном соке растворены сахара (глюкоза, фруктоза, сахароза, инулин), растворимые белки, органические кислоты (щавелевая, яблочная, лимонная, винная, муравьиная, уксусная и др.), разнообразные гликозиды, дубильные вещества, алкалоиды (атропин , папаверин, морфин и др.), ферменты, витамины, фитонциды и др. В клеточном соке многих растений имеются пигменты - антоциан (красный, синий, фиолетовый цвет разных оттенков), антохлоры (желтый цвет), антофеины (темно-бурый цвет). В вакуолях семян содержатся белки-протеины. В клеточном соке растворены также многие неорганические соединения.
Вакуоли - места отложений конечных продуктов обмена веществ.
Вакуоли формируют внутреннюю водную среду клетки, с их помощью осуществляется регуляция водно-солевого обмена. Вакуоли поддерживают тургорное гидростатическое давление внутри клеток, что способствует поддержанию формы неодревесневших частей растений - листьев, цветков. Тургорное давление связано с избирательной проницаемостью тонопласта для воды и явлением осмоса - односторонней диффузией воды через полупроницаемую перегородку в сторону водного раствора солей большей концентрации. Поступающая в клеточный сок вода оказывает давление на цитоплазму, а через нее - на стенку клетки, вызывая упругое ее состояние, т.е. обеспечивая тургор. Нехватка воды в клетке ведет к плазмолизу, т.е. к сокращению объема вакуолей и отделению протопластов от оболочки. Плазмолиз может быть обратимым.
Включения - вещества, образующиеся в результате жизнедеятельности клетки либо про запас, либо как отбросы. Включения локализуются либо в гиалоплазме и органоидах, либо в вакуоле в твердом или жидком состоянии. Включения представляют собой запасные питательные вещества, например, зерна крахмала в клубнях картофеля, луковицах, корневищах и в других органах растений, откладывающиеся в особом типе лейкопластов - амилопластах.
Клеточная стенка - это твердое структурное образование, придающее каждой клетке форму и прочность. Она выполняет защитную роль, предохраняя клетку от деформации, противостоит высокому осмотическому давлению большой центральной вакуоли и препятствует разрыву клетки. Клеточная стенка - продукт жизнедеятельности протопласта. Первичная клеточная стенка образуется сразу после деления клеток и состоит в основном из пектиновых веществ и целлюлозы. Разрастаясь, она округляется, образуя межклетники, заполненные водой, воздухом или пектиновыми веществами. При отмирании протопласта мертвая клетка способна проводить воду и выполнять свою механическую роль.
Клеточная стенка может разрастаться только в толщину. На внутренней поверхности первичной клеточной стенки начинает откладываться вторичная клеточная стенка. Утолщение бывает внутренним и наружным. Наружные утолщения возможны только на свободной поверхности, например, в виде шипов, бугорков и других образований (споры, пыльцевые зерна). Внутреннее утолщение представлено скульптурными утолщениями в виде колец, спиралей, сосудов и т.д. Неутолщенными остаются только поры - места во втроричной стенке клетки. Через поры по плазмодесмам - тяжам цитоплазмы - осуществляется обмен веществ между клетками, передается раздражение из одной клетки в другую и т.д. Поры бывают простые и окаймленные. Простые поры встречаются в паренхимных и прозенхимных клетках, окаймленные - сосудах и трахеидах, проводящих воду и минеральные вещества.
Вторичная клеточная стенка построена главным образом из целлюлозы, или клетчатки (С 6 Н 10 О 5)n - очень стойкого вещества, нерастворимого в воде, кислотах и щелочах.
С возрастом клеточные стенки претерпевают видоизменения, пропитываются различными веществами. Типы видоизменений: опробковение, одревеснение, кутинизация, минерализация и ослизнение. Так, при опробковении клеточные стенки пропитываются особым веществом суберином, при одревеснении - лигнином, при кутинизации - жироподобным веществом кутином, при минерализации - минеральными солями, чаще всего углекислым кальцием и кремнеземом, при ослизнении клеточные стенки поглощают большое количество воды и сильно разбухают.
Ферменты, витамины, фитогормоны. Ферменты - это органические катализаторы белковой природы, присутствуют во всех органоидах и компонентах клетки.
Витамины - органические вещества разного химического состава, присутствуют в качестве компонентов в ферментах и выполняют роль катализаторов. Витамины обозначаются заглавными буквами латинского алфавита: А, В, С, D и др. Различают водорастворимые витамины (В, С, РР, Н и др.) и жирорастворимые (А, D, Е).
Водорастворимые витамины находятся в клеточном соке, а жирорастворимые - в цитоплазме. Известно более 40 витаминов.
Фитогормоны - физиологически активные вещества. Наиболее изучены гормоны роста - ауксин и гиббереллин.
Жгутики и реснички. Жгутики - двигательные приспособления у прокариот и у большинства низших растений.
Реснички имеют многие водоросли, мужские половые клетки высших растений, за исключением покрытосеменных и части голосеменных.
Растительные ткани
1. Общая характеристика и классификация тканей.
2. Образовательные ткани.
3. Покровные ткани.
4. Основные ткани.
5. Механические ткани.
6. Проводящие ткани.
7. Выделительные ткани.
Понятие о тканях как группах сходных клеток появилось уже в трудах первых ботаников-анатомов в XVII в. Мальпиги и Грю описали важнейшие ткани, в частности ввели понятия о паренхиме и прозенхиме.
Классификация тканей на основе физиологических функций была разработана в конце XIX - начале XX в. Швенденером и Габерландтом.
Ткани - это группы клеток, имеющие однородное строение, одинаковое происхождение и выполняющие одну и ту же функцию.
В зависимости от выполняемой функции различают следующие типы тканей: образовательные (меристемы), основные, проводящие, покровные, механические, выделительные. Клетки, составляющие ткань и имеющие более или менее одинаковое строение и функции, называют простыми, если клетки неодинаковые, то ткань называют сложной или комплексной.
Ткани делят на образовательные, или меристемные, и постоянные (покровные, проводящие, основные и т.д.).
Классификация тканей.
1. Образовательные ткани (меристемы):
1) верхушечные;
2) боковые: а) первичные (прокамбий, перицикл);
б) вторичные (камбий, феллоген)
3) вставочные;
4) раневые.
2. Основные:
1) ассимиляционная паренхима;
2) запасающая паренхима.
3. Проводящие:
1) ксилема (древесина);
2) флоэма (луб).
4. Покровные (пограничные):
1) наружные: а) первичные (эпидерма);
б) вторичные (перидерма);
в) третичные (корка, или ритидом)
2) наружные: а) ризодерма;
б) веламен
3) внутренние: а) эндодерма;
б) экзодерма;
в) обкладочные клетки проводящих пучков в листьях
5. Механические (опорные, скелетные) ткани:
1) колленхима;
2) склеренхима:
а) волокна;
б) склереиды
6. Выделительные ткани (секреторные).
2. Образовательные ткани. Образовательные ткани, или меристемы, - это постоянно молодые, активно делящиеся группы клеток. Находятся они в местах роста разных органов: кончиках корней, верхушках стеблей и т.д. Благодаря меристемам происходят рост растения и образование новых постоянных тканей и органов.
В зависимости от местоположения в теле растения образовательная ткань может быть верхушечной, или апикальной, боковой, или латеральной, вставочной, или интеркалярной, и раневой. Образовательные ткани делят на первичные и вторичные. Так, верхушечные меристемы всегда первичные, они определяют рост растения в длину. У низкоорганизованных высших растений (хвощи, некоторые папоротники) верхушечные меристемы слабо выражены и представлены всего лишь одной начальной, или инициальной делящейся клеткой. У голосеменных и покрытосеменных верхушечные меристемы хорошо выражены и представлены многими инициальными клетками, образующими конусы нарастания.
Латеральные меристемы, как правило, вторичны и за счет них происходит разрастание осевых органов (стеблей, корней) в толщину. К боковым меристемам относят камбий и пробковый камбий (феллоген), деятельность которого способствует образованию пробки в корнях и стеблях растения, а также особую ткань проветривания - чечевички. Боковая меристема, как и камбий, образует клетки древесины и луба. В неблагоприятные периоды жизни растения деятельность камбия замедляется или совсем прекращается. Интеркалярные, или вставочные, меристемы чаще всего первичны и сохраняются в виде отдельных участков в зонах активного роста, например у основания междоузлий и у основания черешков листьев злаков.
3. Покровные ткани. Покровные ткани защищают растение от неблагоприятных воздействий внешней среды: солнечного перегрева, излишнего испарения, резкого перепада температуры воздуха, иссушающего ветра, механического воздействия, от проникновения вовнутрь растения болезнетворных грибов и бактерий и т.д. Различают первичную и вторичную покровные ткани. К первичным покровным тканям относятся кожица, или эпидерма, и эпиблема, к вторичным - перидерма (пробка, пробковый камбий и феллодерма).
Кожица, или эпидерма, покрывает все органы однолетних растений, молодые зеленые побеги многолетних древесных растений текущего вегетационного периода, надземные травянистые части растений (листья, стебли и цветки). Эпидерма чаще всего состоит из одного слоя плотно сомкнутых клеток без межклеточного пространства. Она легко снимается и представляет собой тонкую прозрачную пленку. Эпидерма - живая ткань, состоит из постепенного слоя протопласта с лейкопластами и ядром, крупной вакуоли, занимающей почти всю клетку. Стенка клеток в основном целлюлозная. Наружная стенка эпидермальных клеток более утолщенная, боковые и внутренние - тонкие. Боковые и внутренние стенки клеток имеют поры. Основная функция эпидермы - регуляция газообмена и транспирации, осуществляемая в основном через устьица. Вода и неорганические вещества проникают через поры.
Клетки эпидермы разных растений неодинаковы по форме и размерам. У многих однодольных растений клетки вытянуты в длину, у большинства двудольных имеют извилистые боковые стенки, что повышает плотность их сцепления друг с другом. Эпидерма верхней и нижней частей листа также отличается своим строением: на нижней стороне листа в эпидерме большее число устьиц, а на верхней стороне их гораздо меньше; на листьях водных растений с плавающими на поверхности листьями (кубышка, кувшинка) устьица есть только на верхней стороне листа, а у полностью погруженных в воду растений устьица отсутствуют.
Устьица - высокоспециализированные образования эпидермы, состоят из двух замыкающих клеток и щелевидного образования между ними - устьичной щели. Замыкающие клетки, имеющие полулунную форму, регулируют размер устьичной щели; щель может открываться и закрываться в зависимости от тургорного давления в замыкающих клетках, содержания диоксида углерода в атмосфере и других факторов. Так, днем, когда клетки устьиц участвуют в фотосинтезе, тургорное давление в устьичных клетках высокое, устьичная щель открыта, ночью, наоборот, закрыта. Подобное явление наблюдается в засушливое время и при увядании листьев, связано с приспособлением устьиц запасать влагу внутри растения. У многих видов, произрастающих в районах с избыточным увлажнением, особенно во влажных тропических лесах, имеются устьица, через которые выделяется вода. Устьица получили название гидатоды. Вода в виде капель выделяется наружу и капает с листьев. «Плач» растения - своеобразный предсказатель погоды и по научному называется гуттацией. Гидатоды расположены по краю листа, у них нет механизма открывания и закрывания.
В эпидерме многих растений есть защитные приспособления от неблагоприятных условий: волоски, кутикула, восковой налет и др.
Волоски (трихомы) - своеобразные выросты эпидермы, они могут покрывать все растение или некоторые его части. Волоски бывают живыми и мертвыми. Волоски способствуют уменьшению испарения влаги, предохраняют растение от перегрева, поедания животными и от резких колебаний температуры. Поэтому волосками чаще всего покрыты растения аридных - засушливых областей, высокогорий, приполярных районов земного шара, а также растения засоренных местообитаний.
Волоски бывают одноклеточными и многоклеточными. Одноклеточные волоски представлены в виде сосочков. Сососчки встречаются на лепестках многих цветков, придавая им бархатистость (тагетисы, анютины глазки). Одноклеточные волоски могут быть простыми (на нижней стороне многих плодовых культур), и, как правило, они мертвые. Одноклеточные волоски могут быть ветвистые (пастушья сумка). Чаще волоски бывают многоклеточными, различающимися по строению: линейными (листья картофеля), кустисто-ветвистыми (коровяк), чешуйчатыми и звездчато-чешуйчатыми (представители семейства Лоховые), массивными (пучки волосков растений семейства Губоцветные). Встречаются железистые волоски, в которых могут накапливаться эфирные вещества (губоцветные и зонтичные растения), жгучие вещества (крапива) и др. Жгучие волоски крапивы, шипы розы, ежевики, шипы на плодах зонтичных, дурмана, каштана и др. - своеобразные выросты, называемые эмергенцами, в формировании которых принимают участие помимо клеток эпидермы более глубокие слои клеток.
Эпиблема (ризодерма) - первичная однослойная покровная ткань корня. Образуется из наружных клеток апикальной меристемы корня вблизи корневого чехлика. Эпиблема покрывает молодые корневые окончания. Через нее осуществляется водно-минеральное питание растения из почвы. В эпиблеме много митохондрий. Клетки эпиблемы тонкостенны, с более вязкой цитоплазмой, лишены устьиц и кутикулы. Эпиблема недолговечна и постоянно обновляется за счет митотических делений.
Перидерма - сложный многослойный комплекс вторичной покровной ткани (пробка, пробковый камбий, или феллоген, и феллодерма) стеблей и корней многлетних двудольных растений и голосеменных, которые способны непрерывно утолщаться. К осени первого года жизни побеги одревесневают, что заметно по изменению их окраски от зеленой до буро-серой, т.е. произошла смена эпидермы на перидерму, способную выдержать неблагоприятные условия зимнего периода. В основе перидермы лежит вторичная меристема - феллоген (пробковый камбий), образующийся в клетках основной паренхимы, лежащей под эпидермой.
Феллоген образует клетки в двух направлениях: наружу - клетки пробки, внутрь - живые клетки феллодермы. Пробка состоит из отмерших клеток, заполненных воздухом, они вытянуты в длину, плотно прилегают друг к другу, поры отсутствуют, клетки воздухо- и водонепроницаемы. Клетки пробки имеют коричневый или желтоватый цвет, который зависит от присутствия в клетках смолистых или дубильных веществ (пробковый дуб, бархат сахалинский). Пробка хороший изоляционный материал, не проводит тепла, электричества и звуки, используется для закупорки бутылок и др. Мощный слой пробки имеет пробковый дуб, виды бархата, пробковый вяз.
Чечевички - «вентиляционные» отверстия в пробке для обеспечения газо- и водообмена живых, более глубоко лежащих тканей растения с внешней средой. Внешне чечевички похожи на семена чечевицы, за что и получили свое название. Как правило, чечевички закладываются на смену устьицам. Формы и размеры чечевичек различны. В количественном отношении чечевичек намного меньше, чем устьиц. Чечевички представляют собой округлые тонкостенные бесхлорофилльные клетки с межклетниками, которые приподнимают кожицу и разрывают ее. Этот слой рыхлых слабо опробковевших паренхимных клеток, составляющих чечевичку, называется выполняющей тканью.
Корка - мощный покровный комплекс из отмерших наружных клеток перидермы. Она формируется на многолетних побегах и корнях древесных растений. У корки трещиноватая и неровная форма. Она предохраняет стволы деревьев от механических повреждений, низовых пожаров, низких температур, солнечных ожогов, проникновения болезнетворных бактерий и грибов. Растет корка за счет нарастания под ней новых слоев перидермы. У древесно-кустарниковых растений корка возникает (например, у сосны) на 8-10-м, а у дуба - на 25-30-м году жизни. Корка входит в состав коры деревьев. Снаружи она постоянно слущивается, сбрасывая с себя всевозможные споры грибов и лишайников.
4. Основные ткани. Основная ткань, или паренхима, занимает большую часть пространства между другими постоянными тканями стеблей, корней и других органов растений. Основные ткани состоят в основном из живых клеток, разнообразных по форме. Клетки тонкостенные, но иногда утолщенные и одревесневшие, с постенной цитоплазмой, простыми порами. Из паренхимы состоят кора стеблей и корней, сердцевина стеблей, корневищ, мякоть сочных плодов и листьев, она служит хранилищем питательных веществ в семенах. Выделяют несколько подгрупп основных тканей: ассимиляционную, запасающую, водоносную и воздухоносную.
Ассимиляционная ткань, или хлорофиллоносная паренхима, или хлоренхима, - ткань, в которой осуществляется фотосинтез. Клетки тонкостенны, содержат хлоропласты, ядро. Хлоропласты, как и цитоплазма, расположены постенно. Хлоренхима находится непосредственно под кожицей. В основном хлоренхима сосредоточена в листьях и молодых зеленых побегах растений. В листьях различают палисадную, или столбчатую, и губчатую хлоренхиму. Клетки палисадной хлоренхимы удлиненные, цилиндрической формы, с очень узкими межклетниками. Губчатая хлоренхима имеет более или менее округлые рыхло расположенные клетки с большим количеством межклетников, заполненных воздухом.
Аэренхима,или воздухоносная ткань, - паренхима со значительно развитыми межклетниками в разных органах характерна для водных, прибрежно-водных и болотных растений (камыши, ситники, кубышки, рдесты, водокрасы и др.), корни и корневища которых находятся в иле, бедном кислородом. Атмосферный воздух доходит до подводных органов через фотосинтетическую систему посредством передаточныхклеток. Кроме того, воздухоносные межклетники сообщаются с атмосферой с помощью своеобразных пневматод - устьиц листьев и стеблей, пневматод воздушных корней некоторых растений (монстера, филодендрон, фикус баньян и др.), щелей, отверстий, каналов, окруженных клетками-регуляторами сообщений. Аэренхима уменьшает удельный вес растения, что, вероятно, способствует поддержанию вертикального положения водных растений, а водным растениям с плавающими на поверхности воды листьями - удержанию листьев на поверхности воды.
Водоносная тканьзапасает воду в листьях и стеблях суккулентных растений (кактусы, алоэ, агавы, толстянки и др.), а также растений засоленных местообитаний (солерос, биюргун, сарсазан, солянки, гребенщик, черный саксаул и др.), как правило, в аридных областях. Листья злаков также имеют крупные водоносные клетки со слизистыми веществами, удерживающими влагу. Хорошо развитые водоносные клетки имеет мох сфагнум.
Запасающие ткани - ткани, в которых в определенный период развития растения откладывают продукты обмена - белки, углеводы, жиры и др. Клетки запасающей ткани обычно тонкостенны, паренхима живая. Запасающие ткани широко представлены в клубнях, луковицах, утолщенных корнях, сердцевине стеблей, эндосперме и зародышах семян, паренхиме проводящих тканей (фасоль, ароидные), вместилищах смол и эфирных масел в листьях лавра, камфарного дерева и др. Запасающая ткань может превращаться в хлоренхиму, например, при прорастании клубней картофеля, луковиц луковичных растений.
5. Механические ткани. Механические, или опорные, ткани- это своего рода арматура,или стереом. Термин стереом происходит от греческого «стереос» - твердый, прочный. Основная функция - обеспечение сопротивления статическим и динамическим нагрузкам. В соответствии с функциями они имеют подобающее строение. У наземных растений они наиболее развиты в осевой части побега - стебле. Клетки механической ткани могут располагаться в стебле либо по периферии, либо сплошным цилиндром, либо отдельными участками в гранях стебля. В корне, который выдерживает в основном сопротивление на разрыв, механическая ткань сосредоточена в центре. Особенность строения этих клеток - сильное утолщение клеточных стенок, которые и придают тканям прочность. Наиболее хорошо развиты механические ткани у древесных растений. По строению клеток и характеру утолщений клеточных стенок механические ткани разделяют на два типа: колленхиму и склеренхиму.
Колленхима - это простая первичная опорная ткань с живыми содержимым клеток: ядром, цитоплазмой, иногда с хлоропластами, с неравномерно утолщенными клеточными стенками. По характеру утолщений и соединения клеток между собой различают три типа колленхимы: уголковую, пластинчатую и рыхлую. Если клетки утолщены только по углам, то это уголковая колленхима,а если стенки утолщены параллельно поверхности стебля и утолщение равномерное, то это пластинчатая колленхима. Клетки уголковой и пластинчатой колленхимы расположены плотно друг к другу, не образуя межклетников. Рыхлая колленхимаимеет межклетники, а утолщенные клеточные стенки направлены в сторону межклетников.
Эволюционно колленхима возникла из паренхимы. Формируется колленхима из основной меристемы и находится под эпидермой на расстоянии одного или нескольких слоев от нее. В молодых стеблях побегов она располагается в виде цилиндра по периферии, в жилках крупных листьев - по обеим их сторонам. Живые клетки колленхимы способны расти в длину, не препятствуя росту молодых растущих частей растения.
Склеренхима - наиболее распространенная механическая ткань, состоящая из клеток с одревесневшими (за исключением лубяных волокон льна) и равномерно утолщенными клеточными стенками с немногочисленными щелевидными порами. Клетки склеренхимы вытянуты в длину и имеют прозенхимную форму с заостренными концами. Оболочки склеренхимных клеток по прочности близки к стали. Содержание лигнина в этих клетках повышает прочность склеренхимы. Склеренхима есть почти во всех вегетативных органах высших наземных растений. У водных ее либо совсем нет, либо она слабо представлена в погруженных органах водных растений.
Различают первичную и вторичную склеренхиму. Первичная склеренхима происходит из клеток основной меристемы - прокамбия или перицикла, вторичная - из клеток камбия. Различают два типа склеренхимы: склеренхимные волокна, состоящие из мертвых толстостенных клеток с заостренными концами, с одревесневшей оболочкой и немногочисленными порами, как у лубяных и древесинных волокон, или волокон либроформа, и склереиды - структурные элементы механической ткани, располагающиеся в одиночку или группами между живыми клетками разных частей растения: кожуры семян, плодов, листьев, стеблей. Основная функция склереид - противостоять сдавливанию. Форма и размеры склереид разнообразны.
6. Проводящие ткани. Проводящие ткани транспортируют питательные вещества в двух направлениях. Восходящий (транспирационный) токжидкости (водные растворы и соли) идет по сосудам и трахеидамксилемы от корней вверх по стеблю к листьям и другим органам растения. Нисходящий ток (ассимиляционный) органических веществ осуществляется от листьев по стеблю к подземным органам растения по специальным ситовидным трубкамфлоэмы. Проводящая ткань растения чем-то напоминает кровеносную систему человека, так как имеет осевую и радиальную сильно разветвленную сеть; питательные вещества попадают в каждую клеточку живого растения. В каждом органе растения ксилема и флоэма располагаются рядом и представлены в виде тяжей - проводящих пучков.
Существуют первичные и вторичные проводящие ткани. Первичные дифференцируются из прокамбия и закладываются в молодых органах растения, вторичные проводящие ткани более мощные, формируются из камбия.
Ксилема (древесина) представлена трахеидами и трахеями, или сосудами.
Трахеиды - вытянутые замкнутые клетки с косо срезанными зазубренными концами, в зрелом состоянии представлены мертвыми прозенхимными клетками. Длина клеток в среднем 1-4 мм. Сообщение с соседними трахеидами происходит через простые или окаймленные поры. Стенки неравномерно утолщены, по характеру утолщения стенок различают трахеиды кольчатые, спиральные, лестничные, сетчатые и пористые. У пористых трахеид всегда окаймленные поры. Спорофиты всех высших растений имеют трахеиды, а у большинства хвощевидных, плауновидных, папоротниковидных и голосеменных они служат единственными проводящими элементами ксилемы. Трахеиды выполняют две основные функции: проведение воды и механическое укрепление органа.
Трахеи,или сосуды, - главнейшие водопроводящие элементы ксилемы покрытосеменных растений. Трахеи представляют собой полые трубки, состоящие из отдельных члеников; в перегородках между члениками находятся отверстия - перфорации,благодаря которым осуществляется ток жидкости. Трахеи, как и трахеиды, - это замкнутая система: концы каждой трахеи имеют скошенные поперечные стенки с окаймленными порами. Членики трахей крупнее, чем трахеиды: в поперечнике составляют у разных видов растений от 0,1-0,15 до 0,3 - 0,7 мм. Длина трахей от нескольких метров до нескольких десятков метров (у лиан). Трахеи состоят из мертвых клеток, хотя на начальных стадиях формирования они живые. Считают, что трахеи в процессе эволюции возникли из трахеид.
Сосуды и трахеиды помимо первичной оболочки в большинстве имеют вторичные утолщения в виде колец, спиралей, лестниц и т.д. Вторичные утолщения образуются на внутренней стенке сосудов. Так, в кольчатом сосуде внутренние утолщения стенок в виде колец, находящихся на расстоянии друг от друга. Кольца расположены поперек сосуда и чуть наклонно. В спиральном сосуде вторичная оболочка наслаивается изнутри клетки в виде спирали; в сетчатом сосуде неутолщенные места оболочки выглядят в виде щелей, напоминающих ячеи сетки; в лестничном сосуде утолщенные места чередуются с неутолщенными, образуя подобие лестницы.
Трахеиды и сосуды - трахеальные элементы - распределяются в ксилеме различным образом: на поперечном срезе сплошными кольцами, образуя кольцесосудистую древесину, или рассеянно более или менее равномерно по всей ксилеме, образуя рассеянно-сосудистую древесину. Вторичная оболочка, как правило, пропитывается лигнином, придавая растению дополнительную прочность, но в то же время ограничивая его рост в длину.
Помимо сосудов и трахеид ксилема включает лучевые элементы, состоящие из клеток, образующих сердцевинные лучи. Сердцевинные лучи состоят из тонкостенных живых паренхимных клеток, по которым питательные вещества оттекают в горизонтальном направлении. В ксилеме присутствуют также живые клетки древесинной паренхимы, которые функционируют в качестве ближнего транспорта, и служат местом хранения запасных веществ. Все элементы ксилемы происходят из камбия.
Флоэма - проводящая ткань, по которой транспортируется глюкоза и другие органические вещества - продукты фотосинтеза от листьев к местам их использования и отложения (к конусам нарастания, клубням, луковицам, корневищам, корням, плодам, семенам и др.). Флоэма также бывает первичная и вторичная. Первичная флоэма формируется из прокамбия, вторичная (луб) - из камбия. В первичной флоэме отсутствуют сердцевинные лучи и менее мощная система ситовидных элементов, нежели у трахеид.
В процессе формирования ситовидной трубки в протопласте клеток - члеников ситовидной трубки появляются слизевые тельца, принимающие участие в образовании слизевого тяжа около ситовидных пластинок. На этом формирование членика ситовидной трубки заканчивается. Функционируют ситовидные трубки у большинства травянистых растений один вегетационный период и до 3-4 лет у древесно-кустарниковых растений. Ситовидные трубки состоят из ряда удлиненных клеток, сообщающихся друг с другом посредством продырявленных перегородок - ситечек. Оболочки функционирующих ситовидных трубок не одревесневают и остаются живыми. Старые клетки закупориваются так называемым мозолистым телом, а потом отмирают и под давлением на них более молодых функционирующих клеток сплющиваются.
К флоэме относится лубяная паренхима, состоящая из тонкостенных клеток, в которых откладываются запасные питательные вещества. По сердцевинным лучамвторичной флоэмы осуществляется также ближняя транспортировка органических питательных веществ - продуктов фотосинтеза.
Проводящие пучки - тяжи, образуемые, как правило, ксилемой и флоэмой. Если к проводящим пучкам примыкают тяжи механической ткани (чаще склеренхимы), то такие пучки называют сосудисто-волокнистыми. В проводящие пучки могут быть включены и другие ткани - живая паренхима, млечники и др. Проводящие пучки могут быть полными, когда присутствуют и ксилема и флоэма, и неполными, состоящими только из ксилемы (ксилем-ный, или древесинный, проводящий пучок) или флоэмы (флоэмный, или лубяной, проводящий пучок).
Проводящие пучки первоначально образовались из прокамбия. Выделяют несколько типов проводящих пучков. Часть прокамбия может сохраниться и затем превратиться в камбий, тогда пучок способен к вторичному утолщению. Это открытые пучки. Такие проводящие пучки преобладают у большинства двудольных и голосеменных растений. Растения, имеющие открытые пучки, способны разрастаться в толщину за счет деятельности камбия, причем древесинные участки примерно в три раза крупнее лубяных участков. Если при дифференци-ровке проводящего пучка из прокамбиального тяжа вся образовательная ткань полностью расходуется на формирование постоянных тканей, то пучок называется закрытым.
Закрытые проводящие пучки встречаются в стеблях однодольных растений. Древесина и луб в пучках могут иметь различное взаимное расположение. В связи с этим выделяют несколько типов проводящих пучков: коллатеральные, биколлатеральные, концентрические и радиальные. Коллатеральные,или бокобочные,- пучки, в которых ксилема и флоэма примыкают друг к другу. Биколлатеральные,или двубокобочные,- пучки, в которых к ксилеме примыкают бок о бок два тяжа флоэмы. В концентрическихпучках ткань ксилемы полностью окружает ткань флоэмы или наоборот. В первом случае такой пучок называют центрофлоэмным. Центрофлоэмные пучки имеются у стеблей и корневищ некоторых двудольных и однодольных растений (бегония, щавель, ирис, многие осоковые и лилейные).
Ими обладают папоротники. Существуют и промежуточные проводящие пучки между закрытыми коллатеральными и центрофлоэмными. В корнях встречаются радиальныепучки, в которых центральную часть и лучи по радиусам оставляет древесина, причем каждый луч древесины состоит из центральных более крупных сосудов, постепенно уменьшаясь по радиусам. Число лучей у разных растений неодинаково. Между древесинными лучами располагаются лубяные участки. Проводящие пучки тянутся вдоль всего растения в виде тяжей, которые начинаются в корнях и проходят вдоль всего растения по стеблю к листьям и другим органам. В листьях они называются жилками. Главная функция их - проведение нисходящего и восходящего токов воды и питательных веществ.
7. Выделительные ткани. Выделительные, или секреторные, ткани представляют собой специальные структурные образования, способные выделять из растения или изолировать в его тканях продукты метаболизма и капельно-жидкую среду. Продукты метаболизма называют секретами. Если они выделяются наружу, то это ткани наружной секреции, если остаются внутри растения, то - внутренней секреции. Как правило, это живые паренхимные тонкостенные клетки, однако по мере накопления в них секрета они лишаются протопласта и их клетки опробковевают.
Образование жидких секретов связано с деятельностью внутриклеточных мембран и комплекса Гольджи, а их происхождение - с ассимиляционными, запасающими и покровными тканями. Основная функция жидких секретов заключается в защите растения от поедания животными, повреждения насекомыми или болезнетворными микроорганизмами. Ткани внутренней секреции представлены в виде клеток-идиобластов, смоляных ходов, млечников, эфиромасличных каналов, вместилищ выделений, железистых головчатых волосков, железок.В клетках-идиобластах часто содержатся кристаллы щавелевокислого кальция (представители семейства Лилейные, Крапивные и др.), слизи (представители семейства Мальвовые и др.), терпеноиды (представители семейств Магнолиевые, Перечные и др.) и т.п.
Вегетативные органы высших растений
1. Корень и его функции. Метаморфозы корня.
2. Побег и система побегов.
3. Стебель.
К вегетативным органам растений относятся корень, стебель и лист, составляющие тело высших растений. Тело низших растений (водоросли, лишайники) - слоевище, или таллом, не расчленено на вегетативные органы. Тело высших растений имеет сложное морфологическое или анатомическое строение. Оно последовательно усложняется от мохообразных до цветковых растений благодаря все большей расчлененности тела путем образования системы разветвленных осей, что приводит к увеличению общей площади соприкосновения с окружающей средой. У низших растений - это система талломов, или слоевищ, у высших растений - системы побегов и корней.
Тип ветвлений у разных групп растений различный. Выделяют дихотомическое, или вильчатое, ветвление, когда старый конус нарастания разделяется на два новых. Такой тип ветвления встречается у многих водорослей, некоторых печеночных мхов, плаунов, из покрытосеменных - у некоторых пальм. Существуют изотомная и анизотомная системы осей. При изотом-ной системе после прекращения роста верхушки главной оси под нею вырастают две одинаковые боковые ветви, а при анизотомной одна ветвь резко перерастает другую. Наиболее распространен боковойтип ветвления, при котором на главной оси возникают боковыеоси. Такой тип ветвления присущ ряду водорослей, корням и побегам высших растений. Для высших растений выделяют два типа бокового ветвления: моноподиальное и симподиальное.
При моноподиальном ветвленииглавная ось не прекращает роста в длину и образует ниже конуса нарастания боковые побеги, - которые слабее главной оси. Иногда у моноподиально ветвящихся растений встречается ложная дихотомия, когда рост верхушки главной оси прекращается, а под нею образуются две более или менее одинаковые, перерастающие ее боковые ветви, называемые дихазиями (омела, сирень, каштан конский и др.). Моноподиальное ветвление характерно для многих голосеменных и травянистых покрытосеменных растений. Очень распространено симподиальное ветвление, при котором верхушечная почка побега со временем отмирает и начинает усиленно развиваться одна или несколько боковых почек, становящихся «лидерами». Из них образуются боковые побеги, которые защищают побег, прекративший рост.
Усложнение ветвления, начиная от талломов водорослей, вероятно, произошло в связи с выходом растений на сушу, борьбой за выживание в новой воздушной среде. Вначале эти «земноводные» растения прикреплялись к субстрату с помощью тонких корнеобразных нитей - ризоидов,которые впоследствии в связи с усовершенствованием надземной части растения и необходимостью добычи из почвы больших объемов воды и питательных веществ эволюционировали в более совершенный орган - корень. По поводу очередности происхождения листьев или стебля до сих пор нет единого мнения.
Симподиальное ветвление более эволюционно продвинутое и имеет большое биологическое значение. Так, в случае повреждения верхушечной почки роль «лидера» берет на себя боковой побег. Древесно-кустарниковые растения с симподиальным ветвлением переносят обрезку и формирование кроны (сирень, самшит, облепиха и др.).
Корень и корневая система. Морфология корня. Корень основной орган высшего растения.
Основные функции корня сводятся к закреплению растения в почве, активному поглощению из нее воды и минеральных веществ, синтезу важных органических веществ, например гормонов и других физиологически активных веществ, запасанию веществ.
Функции закрепления растения в почве соответствует анатомическое строение корня. У древесных растений корень обладает, с одной стороны, максимальной прочностью, а другой - большой гибкостью. Выполнению функции закрепления способствует целесообразное расположение гистологических структур (например, древесина сконцентрирована в центре корня).
Корень - осевой орган, обычно цилиндрической формы. Растет до тех пор, пока сохраняется верхушечная меристема, покрытая корневым чехликом. На конце корня никогда не образуются листья. Корень ветвится образуя корневую систему.
Совокупность корней одного растения образует корневую систему. В состав корневых систем входят главный корень, боковые и придаточные корни. Главный корень берет начало от зародышевого корешка. От него отходят боковые корни, которые могут ветвится. Корни, берущие начало от наземных частей растения - листа и стебля, называются придаточными. На способность отдельных частей стебля, побега, иногда листа образовывать придаточные корни основано размножение черенками.
Различают два типа корневых систем - стержневую и мочковатую. У стержневой корневой системы четко выделяется главный корень. Такая система свойственна большинству двудольных растений. Мочковатая корневая система состоит из придаточных корней и наблюдается у большинства однодольных.
Микроскопическое строение корня. На продольном разрезе молодого растущего корня можно выделить несколько зон: зону деления, зону роста, зону всасывания и зону проведения. Верхушку корня, где находится конус нарастания, покрывает корневой чехлик. Чехлик защищает ее от повреждения частицами почвы. Клетки корневого чехлика при прохождении корня через почву постоянно слущиваются и отмирают, а на смену им непрерывно формируются новые за счет деления клеток образовательной ткани кончика корня. Это зона деления. Клетки этой зоны интенсивно растут и вытягиваются вдоль оси корня, образуя зону роста. На расстоянии 1-3 мм от кончика корня находится множество корневых волосков (зона всасывания), которые имеют большую поверхность всасывания и поглощают из почвы воду с минеральными веществами. Корневые волоски недолговечны. Каждый из них представляет собой вырост поверхностной клетки корня. Между всасывающим участком и основанием стебля находится зона проведения.
Центр корня занят проводящей тканью, а между ней и кожицей корня развита ткань, состоящая из крупных живых клеток, - паренхима. Вниз по ситовидным трубкам продвигаются растворы органических веществ, необходимые для роста корней, а снизу вверх по сосудам перемещается вода с растворенными в ней минеральными солями.
Вода и минеральные вещества поглощаются корнями растений в значительной мере независимо, и между двумя процессами нет прямой связи. Вода поглощается благодаря силе, которая представляет собой разность между осмотическим и тургорным давлением, т.е. пассивно. Минеральные вещества поглощаются растениями в результате активного всасывания.
Растения способно не только поглощать минеральные соединения из растворов, но и активно растворять нерастворимые в воде химические соединения. Помимо СО 2 растения выделяют ряд органических кислот - лимонную, яблочную, винную и др., которые способствуют растворению труднорастворимых соединений почвы.
Видоизменения корня. Способность корней к видоизменениям в широких пределах - важный фактор в борьбе за существование. В связи с приобретением дополнительной функций, корни видоизменяются. В них могут накапливаться запасные питательные вещества - крахмал, различные сахара и другие вещества. Утолщенные главные корни моркови, свеклы, репы называются корнеплодами.Иногда утолщаются придаточные корни, как у георгина, они называются корневыми клубнями. На строение корней большое влияние оказывают экологические факторы. У ряда тропических древесных растений, обитающих на бедных кислородом почвах, образуются дыхательные корни.
Они развиваются из подземных боковых коней и растут вертикально вверх, поднимаясь над водой или почвой. Их функция заключается в снабжении подземных частей воздухом, чему способствует тонкая кора, многочисленные чечевички и сильно развита система воздухоносных полостей - межклетников. Воздушные корни способны также поглощать влагу из воздуха. Придаточные корни, вырастающие из надземной части стебля, могут играть роль подпорок. Кони-подпорки часто встречаются у тропических деревьев, растущих по берегам морей в зоне прилива. Они обеспечивают устойчивость растений в зыбком грунте. У деревьев тропического дождевого леса нередко боковые корни приобретают досковидную форму. Досковидные корни развиваются обычно при отсутствии стержневого корня и распространяются в поверхностных слоях почвы.
Корни находятся в сложных взаимоотношениях с организмами, обитающими в почве. В тканях корней некоторых растений (боковых, березовых и некоторых других) поселяются почвенные бактерии. Бактерии питаются органическими веществами корня (преимущественно углеродами) и вызывают в местах своего внедрения разрастания паренхимы - так называемые клубеньки. Клубеньковые бактерии - нитрификаторы обладают способностью превращать атмосферный азот в соединения, которые могут усваиваться растением. Такие боковые, как клевер и люцерна, накапливают от 150 до 300 кг азота на гектар. Кроме того, бобовые используют органические вещества тела бактерий на формирование семян и плодов.
Для подавляющего большинства цветковых растений характерны симбиотические взаимоотношения с грибами.
Зона проведения. После отмирания корневых волосков на поверхности корня оказываются клетки наружного слоя коры. К этому времени оболочки этих клеток становятся слабо проницаемыми для воды и для воздуха. Живое содержимое их отмирает. Таким образом, теперь на поверхности корня вместо живых корневых волосков расположены мертвые клетки. Они защищают внутренние части корня от механических повреждений и болезнетворных бактерий. Следовательно, тот участок корня, на котором уже отмерли корневые волоски, нем ожжет всасывать п
Основные отличия растительной клетки от животной
Клетки представляют собой основную структурную единицу как растений, так и животных. И у тех и у других они имеют очень сходное строение, что свидетельствует об их родственном происхождении. Растительная клетка, равно как и животная, имеет следующее строение: оболочка, ядро, цитоплазма, эндоплазматическая сеть, митохондрии, аппарат Гольджи и различные включения. Несмотря на схожесть, они различаются некоторыми компонентами состава, а также способами питания и процессами жизнедеятельности. Растительная клетка отличается наличием пластид (мембранных органоидов). Эти элементы содержатся в хромопластах, хлоропластах и лейкопластах. Для жизнедеятел
ьности клетки важны хлоропласты, содержащие хлорофиллы. В хлоропластах происходит процесс фотосинтеза. В лейкопластах содержатся питательные вещества, поддерживающие жизнедеятельность клетки растения в экстремальных ситуациях. В хромопластах находятся вещества, которые придают определенную окраску листве и стеблям. Растительная клетка имеет жесткую оболочку, состоящую из целлюлозы. После того как рост прекращается, на первичные стенки оболочки накладывается вторичный. Соседние клетки соприкасаются оболочками и создают единую систему оболочек клеток растения. Еще одна особенность - это наличие пор, именуемых плазмодесмами. Благодаря им цитоплазма и мембранные системы связаны напрямую. Вакуоли всегда присутствуют в клетках растений. Именно это включение в цитоплазму реагирует на ввод и вывод воды. У взрослых клеток присутствует центральная вакуоль, в молодых - мелкие вакуолярные пузырьки. Содержимое их включает в себя различные вещества: органические кислоты, соли, ферменты, белки, ионы, пигменты. Все они участ

вуют в метаболизме клетки. Растительная клетка при делении не образует центриолей.
Внутренний контроль над метаболизмом
За жизнедеятельность клетки отвечает ядро. В нем содержится генетический материал - ДНК, происходит синтез РНК и рибосом. Хроматин, связанный с ДНК, отвечает за синтез белка. Строение цитоплазмы на первый взгляд достаточно простое - вода, органоиды и растворенные вещества. Именно в ней протекают почти все процессы метаболизма клетки. Вся цитоплазма пронизана белковыми нитями и трубочками, которые постоянно образовываются и распадаются под влиянием различных веществ.

Рибосомы, получившие сигнал из ядра на образование новых белковых молекул взамен старых, синтезируют их из растворенных в цитоплазме веществ. Растительная клетка, как и животная, подчиняется информации, заключенной в ДНК. Ядро тоже имеет свою оболочку и поры, при помощи которых происходит передача сигналов для начала тех или иных процессов. Не стоит забывать и о такой важной составляющей, как АТФ. Именно благодаря ему переносятся по цитоплазме вещества для функционирования клетки, происходит избавление от мертвых и ненужных компонентов. К тому же АТФ не только переносит информационные сигналы о начале того или иного процесса, он является поставщиком энергии для клеток.